脂质的消化过程始于唾液分泌、咀嚼以及舌脂酶在口腔中的释放。当消化物进入胃部时,舌脂酶从舌部浆液腺释放后,会通过水解甘油三酯结构中sn-3位点的游离脂肪酸来完成后续分解,其中“sn”表示甘油骨架的立体化学编号方式。进入胃部后,胃脂肪酶继续水解脂质,主要释放出原本作为甘油三酯成分的短链脂肪酸。尽管这两种脂肪酶都参与了水解作用,但进入十二指肠上段的脂质中仍有超过70%是甘油三酯。因此,小肠才是脂质消化的主要场所。

小肠中脂质的消化过程主要依赖两大关键成分:胆汁酸和胰脂肪酶。胆汁酸由肝脏中的胆固醇转化生成,随后被浓缩储存于胆囊。当循环系统中胆囊收缩素(一种肽类激素)水平升高时,就会引发水油乳化现象,从而促使胆汁酸释放至肠道腔内。尽管胆汁酸对于微胶束的形成至关重要,但当其被释放入肠腔时,最初会抑制胰腺脂肪酶的活性。这种抑制是由于胆汁酸物理性地阻止胰腺脂肪酶与肠腔中的脂滴接触。共脂肪酶通过结合胰腺脂肪酶来逆转胆汁酸的抑制作用,两者一旦结合后便能附着于脂滴表面。当胰腺脂肪酶通过共脂肪酶的结合作用附着于脂滴后,便会酶促催化甘油三酯中sn-1和sn-3位的酯键断裂。该酶促水解反应生成两个游离脂肪酸以及一个在sn-2位酯化脂肪酸的单甘酯。此酶促反应速率极快,其产生游离脂肪酸和单甘酯的速度远快于后续微胶束的整合速度。而抗胰腺脂肪酶水解的磷脂则通过磷脂酶A2进行消化。磷脂酶A2通过酶促反应从sn-2位释放脂肪酸,产生溶血磷脂和游离脂肪酸。共脂肪酶将新水解的产物从肠腔中的脂滴运输至正在形成的含胆汁酸盐的微胶束中。
胆汁中的脂质物质会形成胶束结构。这种结构的形成源于胆囊分泌的胆汁酸与磷脂共同作用:胆汁酸的一端朝向消化物和肠腔的水环境,另一端则指向胶束中心。通过这种特殊取向,胆汁酸与磷脂在胶束中形成了疏水核心和亲水边缘的结构特征。研究表明,当脂质被整合到胶束结构时,其对多不饱和脂肪酸(PUFA)和饱和单酰基甘油酯表现出更强的结合能力。混合胶束一旦形成,就会穿过肠腔直达肠上皮细胞的静水层。这种胶束结构巧妙解决了脂质在肠道水环境中易受疏水性影响的问题,不仅让脂质能轻松穿透静水层,还能使肠上皮吸收表面附近的游离脂肪酸、单甘酯等脂质物质浓度提升100至1000倍。图1简明展示了脂质消化与吸收的完整过程。
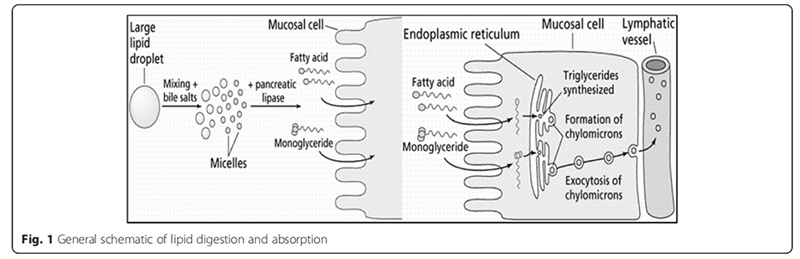
由于胶束中脂质物质的富集形成了浓度梯度,脂质成分可通过非能量依赖的过程被动扩散进入肠细胞。当肠腔内脂质含量较低时,也有证据表明存在依赖载体的脂质双层吸收过程。这种双重机制在理论上被提出用于维持膳食脂质摄入量低时必需脂肪酸的必要水平,但尚不清楚当膳食脂质摄入正常或较高时载体介导的运输是否重要。由于肠道的搅动作用和结构特征,胶束之间保持着动态平衡关系,这使得上皮细胞、胶束和脂滴几乎持续接触。这种高度接触将脂质成分从高密度胶束分配到低密度胶束。这种分配使胶束均匀获取并分布脂质成分,最终意味着小肠腔内脂质消化的限制因素是胶束饱和度。脂质成分从胶束穿越静水层的穿梭过程是一种链式反应,其依赖于肠细胞处脂质的低浓度。肠道脂肪酸结合蛋白通过结合游离脂肪酸,并将其截留在顶膜区域,从而促进脂肪酸的吸收。胆汁酸可在回肠末端被高效重吸收,继而转运至肝脏循环利用,参与后续的脂质消化过程。
当脂肪酸(FA)扩散至肠细胞后,会通过甘油-3-磷酸途径或单酰基甘油途径在内质网中重新酯化。经过甘油三酯的再酯化过程后,多个甘油三酯和胆固醇酯会被包装成乳糜微粒。乳糜微粒含有80%至95 %的甘油三酯、2%至7%的胆固醇以及3%至9%的磷脂。其外层由磷脂双分子层和载脂蛋白构成,这些成分能增强溶解度并提高酶识别能力。随后,乳糜微粒通过胸导管进入淋巴系统,最终进入血液循环系统。
乳糜微粒进入血液循环后,既可储存在脂肪细胞中,也可被肌纤维等细胞氧化分解。当胰岛素和合成代谢激素水平升高时,这些微粒就会被转运至脂肪细胞进行储存。这一过程主要受胰岛素对脂肪细胞脂蛋白脂肪酶的刺激作用调控,而肌肉细胞中的脂蛋白脂肪酶同工酶则不受胰岛素影响。因此,多功能酶脂蛋白脂肪酶会在脂肪细胞的毛细血管腔内表达,专门负责处理富含甘油三酯的乳糜微粒及其他脂蛋白。脂肪酸会以被动扩散方式逐个进入细胞,随后在脂肪细胞内重新酯化形成甘油三酯储存。
与含有16至20个碳原子脂肪酸的长链甘油三酯不同,中链甘油三酯主要由含有8和10个碳原子的饱和脂肪酸构成。当这些脂肪酸被脂肪酶快速分解后,即使在少量腔内胆汁酸和胰脂肪酶存在的情况下,它们仍具有高水溶性,能迅速被黏膜细胞吸收形成乳糜微粒。随后,这些中链脂肪酸与白蛋白结合,通过门静脉系统运输至肝脏,并以不依赖肉碱的方式进入线粒体进行后续氧化反应。
 0536-2096656 0536-2096659
0536-2096656 0536-2096659 info@kenonbio.com
info@kenonbio.com 山东省潍坊市滨海新区先进制造业产业园珠江西二街00508号 0532-58558788 info@kenonbio.com 中国·青岛 崂山区深圳路101号华润置地大厦20层
山东省潍坊市滨海新区先进制造业产业园珠江西二街00508号 0532-58558788 info@kenonbio.com 中国·青岛 崂山区深圳路101号华润置地大厦20层
 EN
EN