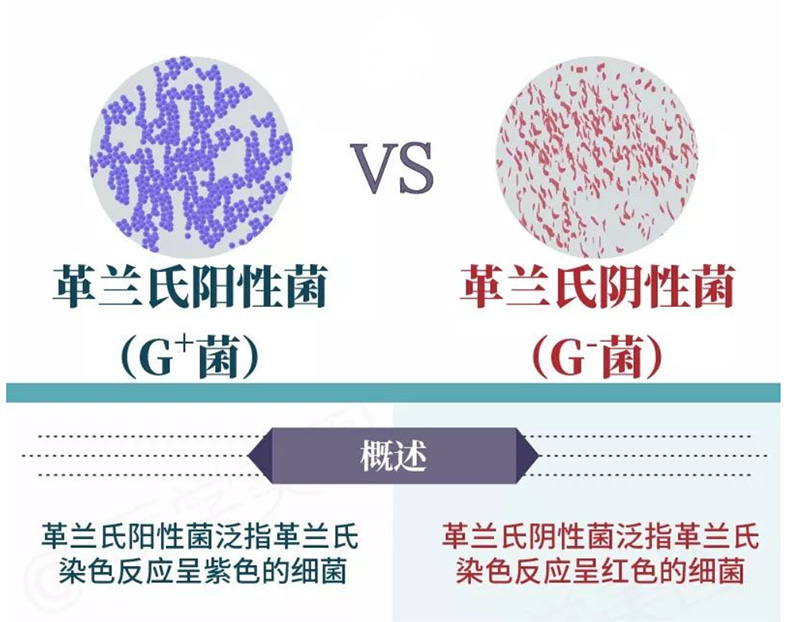
图1.革兰氏阳性菌(左)和革兰氏阴性菌(右)概述
1.金黄色葡萄球菌(Staphylococcus aureus)
研究表明, 破坏生物膜功能和生物能传输过程可能是脱氧胆酸盐针对金黄色葡萄球菌的抑菌机制。最低抑菌浓度下的脱氧胆酸盐处理后, 金黄色葡萄球菌细胞内钾离子渗透性升高, 细胞内pH降低, 细胞膜内外两侧的电势差被破坏, 表明脱氧胆酸盐能阻断生物能传输过程; 同时, 共聚焦显微镜显示脱氧胆酸盐引起了金黄色葡萄球菌细胞膜的严重破坏, 扫描电子显微镜和透射电子显微镜结果显示脱氧胆酸盐处理后的金黄色葡萄球菌细胞形态皱缩, 表面出现凹陷, 细胞壁变薄甚至是破裂, 细胞膜发生破裂, 揭示了脱氧胆酸盐能通过破坏金黄色葡萄球菌的基本膜功能。脱氧胆酸盐引起的细胞壁/膜破损可能是导致细胞内pH下降、离子渗透性增加和膜电位耗散的主要原因。
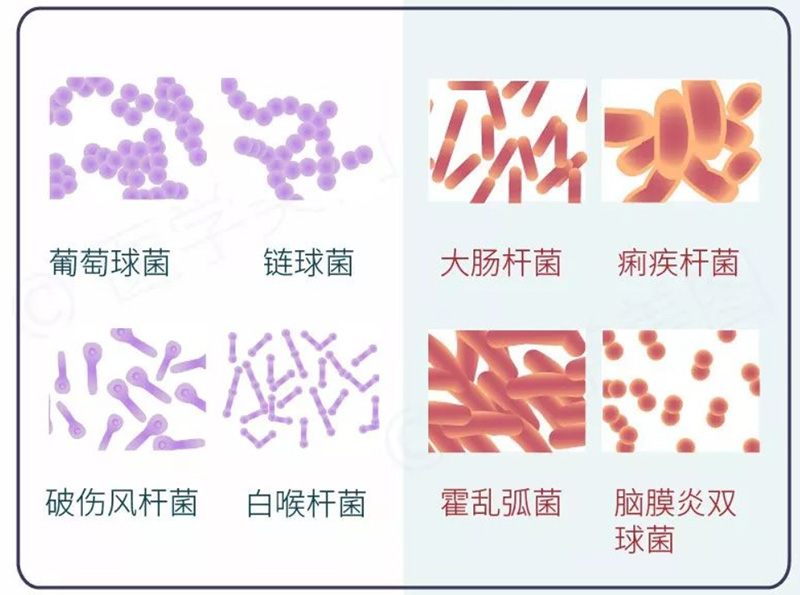
图2.革兰氏阳性菌(左)和革兰氏阴性菌(右)菌种代表
2.艰难梭菌(Clostridioides difficile)
鞭毛作为艰难梭菌的主要运动器官, 对艰难梭菌的致病性有着重要作用. 鞭毛介导艰难梭菌在肠上皮细胞的黏附过程, 是艰难梭菌在肠道中定植的关键步骤。 Metzendorf等人 在扫描显微镜下观察到, 使用生长抑制浓度的DCA和CDCA处理艰难梭菌会导致鞭毛数量减少, 而石胆酸(lithocholic acid, LCA)则会导致鞭毛几乎完全消失。鞭毛中具有代表性鞭毛蛋白(flagellin, FliC)和鞭毛丝帽蛋白(filament cap protein, FliD)丰度较低, 表明LCA、DCA和CDCA应激期间鞭毛合成减少, 鞭毛蛋白合成被抑制。在转录水平上, fliCmRNA丰度与蛋白质水平一样, 在LCA、DCA处理后显著降低, 说明鞭毛蛋白的较低表达已经在转录水平上实现。
3.丙酸杆菌(Propionibacterium)
丙酸杆菌经脱氧胆酸盐/胆酸盐混合刺激后, 其典型多形棒状超微结构发生显著畸变。 扫描电子显微镜显示菌体收缩塌陷, 部分呈现空泡化结构, 收集胞外蛋白进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳, 结果显示胞外蛋白浓度显著升高. 以上结果表明, 脱氧胆酸、胆酸盐混合物破坏丙酸杆菌的细胞壁和细胞膜,并引起胞质溶质外溢。 质谱蛋白质组学结果显示, 与超氧化物歧化酶同源的胆汁盐特异性蛋白B8(bile-saltsspecific proteins B8)的水平显著升高, 提示胆盐对丙酸杆菌造成了氧化损伤, 因为超氧化物歧化酶参与氧化应激引起的损伤修复反应。而在另一项研究中, 利用二维电泳研究了丙酸杆菌SI41暴露于胆盐后细胞中的蛋白质合成, 发现胆盐应激蛋白17(bile salts stress proteins 17, BSSP 17)、胆盐应激蛋白10(bile salts stress proteins 10, BSSP 10) 的蛋白水平显著升高。 BSSP 17被多种方法证明是丙酸杆菌超氧化物歧化酶A, 同时,上调的BSSP 10被鉴定为半胱氨酸合成酶, 半胱氨酸的 合成增强有助于谷胱甘肽的产生, 用于氧化应激期间的解毒。以上实验结果表明, 胆盐不仅会破坏丙酸杆菌表面的细胞膜, 而且会引起氧化应激。
4.咽峡炎链球菌(Streptococcus anginosus)
我们实验室的研究结果表明,12-酮基石胆酸(12-ketodeoxycholic acid, 12-KetoLCA)能显著抑制咽峡炎链球菌的生长和生物膜形成。 最低抑菌浓度的12-KetoLCA处理后, 咽峡炎链球菌胞内蛋白质和核酸发生渗漏, 共聚焦显微镜结果显示12-KetoLCA引起了细胞膜的严重损坏。扫描电子显微镜结果显示, 12-KetoLCA处理后的咽峡炎链球菌细胞明显凹陷变形, 甚至于细胞膜、细胞壁的严重破损, 而在透射电子显微镜下观察到细胞膜发生严重破损, 内容物大量泄漏. 我们同时进行转录组分析, 发现咽峡炎链球菌的基因表达受到12-KetoLCA影响, 包括核糖体蛋白基因的下调, 例如30S核糖体蛋白S12和50S核糖体蛋白L17, 这表明蛋白质合成受损。此外, 我们还观察到参与膜功能的营养转运蛋白的表达模式发生变化. 具体而言, ABC转运蛋白系统基因表达上调, 而与甘露糖、果糖和山梨糖转运相关的磷酸转移酶系统表达下调, 与氨基酸、核苷的转运以相关的表达显著上调。以上结果突显了在12-KetoLCA造成的损伤下膜功能的紊乱。
原文:管健,王玉桂,丁昊,等.胆汁酸对肠道微生态的调控研究进展[J/OL].科学通报,1-9[2026-01-07].https://link.cnki.net/urlid/11.1784.N.20251226.0953.008..
备注:转载只为分享,以学习交流为目的,如有侵权,联系删除。
 0536-2096656 0536-2096659
0536-2096656 0536-2096659 info@kenonbio.com
info@kenonbio.com 山东省潍坊市滨海新区先进制造业产业园珠江西二街00508号 0532-58558788 info@kenonbio.com 中国·青岛 崂山区深圳路101号华润置地大厦20层
山东省潍坊市滨海新区先进制造业产业园珠江西二街00508号 0532-58558788 info@kenonbio.com 中国·青岛 崂山区深圳路101号华润置地大厦20层
 EN
EN